

 The Energy for Muscular Contractions
The Energy for Muscular Contractions
There is only one source of energy for muscular contractions: ATP. Energy, to power muscular contraction, is released when ATP is broken down toadenosine diphosphate (ADP) and phosphate (Pi). The body has several different paths by which it produces ATP; and it is vitally important for the body to have these ATP production mechanisms because only a very small amount of ATP can be stored in the muscle (enough for only a few seconds of maximum muscular effort). For this reason, ATP must be supplied to the muscles on a continuous basis during muscular exertion. There are three ways by which the body produces ATP, all of which take place predominantly at the mitochondria.They are:
- by the phosphagen system – the chemical breakdown of phosphocreatine (PC) – an anaerobic mechansim
- by anaerobic glycolysis – an anaerobic mechanism
- by oxidative phosphorylation (Kreb’s cycle and electron transport) – anaerobic mechanism
The phosphagen system: PC is stored in the muscle. A rested muscle contains ~5 times as much PC as it does actual ATP. When PC is broken down, the energy released is used to recombine Pi and ADP to form ATP again. This process can happen in merely a fraction of a second, and so provides a source of quickly replenishable energy. Because of the nature of this mechanism, ATP stores remain fairly constant during the early stages of muscular contraction, but PC stores get depleted. As contraction continues, there is not adequate CP left to continue fueling the necessary ADP -> ATP conversion, leading to the depletion of ATP stores also. This contributes to fatigue of the fiber. In total, within 30 seconds or less of maximum muscular contraction the ATP and PC stores in the muscle are exhausted. It should be noted that the phosphagen system provides the largest power source of any muscular energy mechanism.

The chemical breakdown of PC cycle
As you can probably figure out, the phosphagen system is the primary energysource for short-term, high-intensity work, such as heavy weighttraining.
NOTE: PC stores cannot be used to provide energy for long-term, low-intensity work.
Anaerobic glycolysis: Glycogen (the form of glucose that is stored in muscle) is broken down to provide the energy for ATP formation and and also the formation of pyruvic acid. Additionally, some blood glucose may be used in this process, along with the intramuscular glycogen. One of the end products of this mechanism is lactic acid, which is made by the eventual conversion of pyruvic acid.
This mechanism can provide more total energy than the phosphagen system, but not as quickly. This being the case, anaerobic glycolysis is the major energy pathway for muscular contractions lasting from ~30 to ~60 seconds.
The effects that the lactic acid (which is produced during this process) has on muscular contraction must be considered here. Lactic acid build-up in the muscle cells make the interior of the muscle more acidic. This acidic environment interferes with the chemical processes that expose actin cross-bridging sites and permit cross-bridging. It also interferes with ATP formation. So, these factors, along with depleted energy stores, contribute to muscle fiber fatigue. Contrary to what was once believed, lactic acid does not cause delayed onset muscle soreness (the soreness that you feel in an exercised muscle the next day or so). High lactic acid concentration does, however, contribute to the sensation of pain in motor nerve endings during muscular contraction.
Oxidative phosphorylation: In this mechanism the body metabolizes carbohydrates and fats (and protein when under starvation conditions or duringvery long duration exercise sessions) to create energy. Carbohydrates are used more extensively during intense aerobic work (at near 100% capacity levelscarbohydrates are used almost exclusively as the energy source) and fats become the primary energy source during low-intensity, long duration exercise sessions. The process of energy release from these substrates is much more complex than we need to get into, and as this process is the least crucial for most weight training activities, it will suffice to say that they require oxygen. Hence, your breathing rate increases during aerobics.
This mechanism provides virtually endless amounts of energy (well, until you collapse) as your body will actually begin to cannibalize itself in order tokeep the process going. It does, however, require time and so is not a major player in supplying energy for intense muscular contractions.
As was mentioned above, certain fiber types are optimized to utilize each of these energy production mechanisms.
Type Is (slow twitch (ST), slow oxidative – called red fibers) utilize primarily the oxidative phosphorylation mechanism.
Type IIAs (fast twitch (FT), fast oxidative – called white fibers) utilize both the phosphagen system and the anaerobic glycolysis mechanismsprimarily.
Type IIBs (fast-glycolytic – a kind of white FT fiber) utilizeprimarily the phosphagen system.
It should also be noted that all three of these mechanisms begin at the start of muscular contraction, but because of their natures, and the natures of the muscle fibers being used for the activity, they only become prominent during the time frames given above. This is illustrated in the graph below.

Time Course ofContributions from Different Energy Sources
Taken fromGleim, Anaerobic Testing and Evaluation, Med Exerc Nutr Health1993;2:27-35
Oxygen is not only used during the processes of oxidative phosphorylation. It is also required in mechanisms which replenish ATP, PC and glycogen. This is one of the reasons why, even if you only do lows reps, you breath heavy between sets of Squats (or any other exercise that utilizes a lot of muscle mass). ATP replenishment occurs roughly in the time frames presented below.
| |
|
| |
|
| |
|
| |
|
| |
|
| |
|
| |
|
| |
|
| |
|
These times assume that the fibers recovering are at rest. If you do anything during this period that depletes ATP then the process would be impaired and the time needed for replenishment lengthened.
If the activity that the muscles were doing generated a lot of lactic acid (anaerobic glycolysis mechanism) – such as intense weight training in the 12 repand above range – then light activity of the muscles during the replenishment period may actually be of benefit. This is because some of the lactic acid wouldbe used to fuel the light activity and, hence, the activity would help clear lactic acid from the muscle. Care must be taken, though, to ensure that thislight activity is not intense enough to require the use of the phosphagen or anaerobic glycolysis mechanisms for energy – this would deplete ATP as warned ofabove.
Once glycogen stores in the muscle are depleted (from prolonged anaerobic glycolysis) they may take several days to be restored. As this is getting intothe realm of nurition, the subtleties of the practice of replenishing glycogenbetween weight training sessions will be covered in an article on the ‘Nutrition And Supplementation Articles’ page.
Muscle fibers are stimulated by the nervous system by way of alpha motor neurons. Each neuron may control only several muscle fibers or as many as athousand or more. Each muscle fiber, however, is innervated by only one neuron. A neuron and the fibers it innervates are referred to as a motor unit. All ofthe muscle fibers in a motor unit (stimulated by the same neuron) tend to be of the same fiber type (more on fiber types later). You may have heard of the ‘all-or-none’ theory in regards to this subject. It states that all of the fibers in a motor unit must fire or none of them, although this may not be 100% true in certain cases (such as fatigue).
How does the neuron ‘innervate’ it’s associated muscle fibers? Well, the neuron ‘connects’ to the fibers at their center (their length-wise center). To innervate them they transmit an electric current – called the ‘action potential’ – to the fibers, which travels out from the center of the fibers to their ends, thus setting off a contraction…
So how does all this happen? Let’s take a closer look.
Anatomy of a Neuron
All nerve cells (called ‘neurons’) outside the central nervous system (the brain and spinal cord) are made up of large cell bodies and single, elongated extensions (called ‘axons’), for sending messages. Many neurons within the central nervous system also have this configuration. At the end of these axons are ‘axon terminals’ which are the point of release of chemicals that transmit impulses across to other cells (i.e. other neurons or muscle cells). Motor neurons connect your spinal cord to your muscles and can, therefore, have very long axons (as much as 1 m long and only a few micrometers in diameter). There is a steady transport of materials (e.g. vesicles, mitochondria, etc) from the ‘cell body’ (which houses the nucleus as well as other organelles) along the entire length of the axon to the axon terminals.
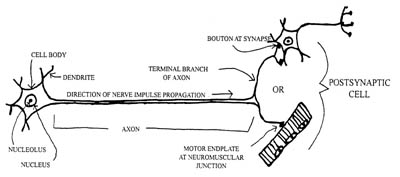
In many neurons, nerve impulses are generated in short branched fibers called ‘dendrites’ and also in the cell body. These impulses are then conducted along the axon, which usually branches several times close to its end for the purpose of innervating several other cells.
The Resting Potential
All cells (not just the neurons) have a resting potential – an electrical charge across their surface membranes (called the ‘plasma membrane’). To produce this the interior of the cell is maintained with a negative charge with respect to the exterior. The size of this resting potential varies with cell type, but in neurons it is about -70 milliVolts (mV) and about -95 mV in muscle cells.
The resting potential is generated and maintained in two ways:
1. The Sodium/Potassium ATPase Pump: There is, typically, a 20 times higher concentration of positively charged potassium ions (K+) inside the cell than outside the cell (in the extracellular fluid). Conversely, the extracellular fluid contains a concentration of positively charged sodium ions (Na+) as much as 10 times greater than that within the cell. These concentration gradients are maintained by the active transport of both ions back and forth across the plasma membrane by the Na+/K+ ATPase transporter system. It transports 3 Na+ ions out of the cellfor each 2 K+ ions pumped in (using energy produced from the breakdown of ATP to fuel the process).
As an aside: Besides just maintaining the cell’s resting potential, this Na+/K+ balance has another function – of interest primarily to Bodybuilders. The accumulation of sodium ions outside of the cell draws water out of the cell and thus enables it to maintain osmotic balance. This is why excessively high sodium levels in the blood make you hold water and look ‘smooth’. Potassium has the opposite effect, so the two are often manipulated by Bodybuilders prior to physique competitions.
2. Facilitated diffusion of K+ out of the cell: Some potassium channels in the plasma membrane are ‘leaky’, allowing a slow diffusion of K+ out of the cell.
Depolarization and the Action Potential
Certain stimuli can cause the Na+/K+ balance across the plasma membrane to change. By far, the most significant of these stimuli are ‘neurotransmitters’ (chemicals which transmit neural stimulation across the gap between neurons and other excitable cells) such as acetylcholine (ACh). These neurotransmitters cause Na+ channels on the plasma membrane to open and Na+ to ‘rush’ into the cell. This, in turn, causes the electric potential across the plasma membrane to decrease, and if it decreases enough (i.e. reaches the ‘threshold voltage’) an ‘action potential’ is generated in the cell. Electrically, this changing of the cell’s resting potential is called ‘depolarization’.
It should be mentioned that certain mechanical stimuli, such as stretching, can also cause Na+ channels to open, thereby setting off an action potential. This helps form the basis (along with some other factors) of what is often called the ‘stretch reflex’ or ‘myotatic reflex’ in muscular contraction. Some strength training authors recommend exploiting this reflex to recruit ‘more muscle fibers’ – this will be examined in an article on the ‘Training Related Articles’ page of The WeighTrainer.
The Action Potential: If depolarization at a spot on the cell reaches the threshold voltage hundreds of sodium channels open in that portion of the plasma membrane. And, even though the channels only remain open for a millisecond (the enzyme acetylcholinesterasequickly breaks down the ACh in the neuromuscular junction, thus allowing the Na+ channels to close again), thousands of Na+ ions rush into the cell. This sudden complete depolarization of the plasma membrane opens up the voltage-gated sodium channels in adjacent portions of the membrane and a ‘wave’ of depolarization sweeps along the cell. This, in fact, is what is called the ‘action potential’ (in neurons it may also be called the ‘nerve impulse’).
The Refractory Period: Another stimulus applied to a neuron (or muscle fiber) cannot trigger another impulse until a sufficient time has passed so that the resting potential can be restored in the plasma membrane. During that ‘refractory’ period the membrane is depolarized and the Na+/K+ ATPase Pump works to restore the Na+/K+ charge balance. This repolarization processes is initiated by the facilitated diffusion of K+ ions out of the cell. Then, when the neuron is fully rested, the sodium ions that came in during the impulse are actively transported back out of the cell.
As was eluded to in Part I of this series, this process of depolarization and repolarization can occur much more rapidly in type II fibers than in type I fibers – leading to a much faster twitch rate in the former. In essence, this is why type II fibers are often referred to as ‘fast twitch’ and type Is as ‘slow twitch’.
Each cell type has only one ‘strength’ of action potential. This means that as long as the threshold potential of the cell is reached, ‘strong’ stimuli will produce no stronger action potentials than ‘weak’ ones. This is what is referred to as the ‘all-or-none’ principle (and, no, I don’t believe Weider has grabbed that one yet). The difference in stimuli strength is reflected by the frequency of the action potentials that it generates. This explains why fractional increases in muscular tension requirements are met by the muscles twitching faster (as was covered in Part I of this series).Skeletal Muscle Motor Neurons Are ‘Myelinated’
The axons of skeletal muscle motor neurons are encased in a fatty sheath called the ‘myelin sheath’ (it is actually the greatly expanded plasma membrane of an accessory cell called the ‘Schwann cell’). Where the sheath of one Schwann cell meets the next, the axon is unprotected. The voltage-gated sodium channels of myelinated neurons are confined to these spots (called ‘nodes of Ranvier’).


The inrush of sodium ions at one node creates just enough depolarization to reach the threshold of the next. In this way, the action potential jumps from one node to the next. This results in much faster propagation of the nerve impulse than is possible in nonmyelinated neurons.
Other Factors
There are other ions that can influence plasma membrane charge balance (most notably chloride – Cl-) and, therefore, affect resting and action potential. Certain neurotransmitters actually inhibit the transmission of nerve impulses by opening chloride ion channels that allow these negatively charged ions to enter the cell. These neurotransmitters may also open K+ channels, allowing potassium ions to ‘leak’ out. The overall result is a state of enhanced plasma polarization called ‘hyperpolarization’. In this state the action potential is ‘further away’ from the resting potential because the resting potential has increased, thus a stronger stimulus is needed to reach the threshold.
The Synapse
Since the junction between the axon terminals of a neuron and other receiving cells (i.e. muscle cells or other nerve cells) is of such importance for transmission of impulses, let’s take a closer look at that junction – called the ‘synapse’. For future possible reference, synapses at muscle fibers are called ‘neuromuscular junctions’ or ‘myoneural junctions’.
src=”images/neuromuscular-junction.gif” ALIGN=RIGHT>
Each axon terminal is swollen into a knob containing membrane-bounded ‘vesicles’ which store neurotransmitters. When an action potential arrives calcium ion (Ca++) channels open in the plasma membrane and trigger some of the vesicles to fuse with the outer cell wall and release their neurotransmitter into the synaptic cleft. These neurotransmitter molecules then bind to receptors on the postsynaptic membrane (which could be the plasma membrane of a muscle cell, for instance), thereby setting of a process of Na+/K+ diffusion and depolarization of the postsynaptic membrane. For a muscle cell this would result in contraction. It should also be mentioned here that the terminal vesicles of motor neurons always cantain the neurotransmitter acetylcholine (ACh).
Ship to Shore: From Excitation to Contraction
In resting muscle fibers, an intracellular organelle called the ‘sarcoplasmic reticulum’ stores calcium ions (Ca++). Spaced along the plasma membrane of the muscle fiber (called the ‘sarcolemma’) are depressions in the membrane that ‘plunge’ into the muscle cell called ‘T-tubules’. These T-tubules (collectively called the ‘T System’) terminate near the calcium-filled sacs of the sarcoplasmic reticulum. Each action potential created at the neuromuscular junction travels along the sarcolemma, down into the T-tubules and innervates the sarcoplasmic reticuli – thus triggering them to release their Ca++ into the interior of the cell. The Ca++ then diffuses among the actin and myosin filaments of the sarcomeres where it binds to the protein troponin. This is of extreme importance in creating a muscular contraction because, under resting conditions, there is a troponin-tropomyosin (a special protein complex) barrier that ‘covers’ the cross-bridge sites (by binding to actin) thus preventing contraction from taking place. Ca++ changes the shape of this troponin-tropomyosin barrier, thereby allowing for cross-bridges to be formed. Without this action the myosin cross-bridges would not be able to make binding contact on the actin filaments. In this way, Ca++ plays the active role in muscle contraction because it ‘turns on’ the interaction between actin and myosin.
Because of the speed of the action potential (milliseconds), the action potential arrives virtually simultaneously at the ends of all the tubules of the T system, ensuring that all sarcomeres contract in unison. When the process is over, the calcium is taken back into the sarcoplasmic reticulum by way of what is called the Ca++/ATPase Pump (or the Ca-Pump).
Conclusion
If you made it through you’ve probably learned a few things that youcan put in the context of your own training already (unless you already knewthis stuff). If not, then this stuff will be referenced heavily in otherarticles on the ‘Training Related Articles’ and ‘Nutrition And SupplementationArticles’ pages. If you really don’t take to the scientific side of weighttraining don’t worry, you didn’t read all that scientific mumbo-jumbo fornothing – it’ll be used in other articles to put together, and make sense of,weight training and nutrition and supplementation practices.
NOTE: The information that has been presented here is by no means extensive.What I have tried to do is present what is relevant to weight training from a weight trainingperspective. Also, it should be realized that the sum of our knowledge today ishopefully smaller than what the sum of our knowledge will be tomorrow, so newfacts and understandings may come along that shed a whole new light on things.If you’re interested, the links below provide more information along theselines.
Please read: The Neuromuscular System Part I
* This article is exclusive to IronMagazine.com, reproduction in any form without prior consent is strictly prohibited.
Visit The WeighTrainer
“Where Training Science Meets Training Reality”


COMMENTS